饲粮中NFC与NDF比例影响肉牛肌肉氨基酸和蛋白质代谢

相关热词搜索:
朱晋佳,吴祎程,吕小康,周传社
(中国科学院 亚热带农业生态研究所 亚热带农业生态过程重点实验室,湖南 长沙 410125)
近年来,为了提高反刍动物的生产性能和动物生产的经济效益,对反刍动物进行了高谷物饲粮的饲喂[1]。适宜的精粗比能够提高营养物质的消化率,最终提高生产性能。改变饲粮精粗比其本质上是改变了饲粮的非纤维碳水化合物(NFC)/中性洗涤纤维(NDF)比例。NFC主要包括淀粉、糖、果胶、果聚糖和有机酸等,是反刍动物的主要能量物质;
NDF是粗饲料的主要组成成分[2]。因此饲粮结构的变化是反刍动物生产中的关键营养调控手段[3]。据报道,提高饲粮NFC/NDF比例能够提高反刍动物的生产性能,这也是生产中通常采用的做法。犊牛在饲喂NFC/NDF为1.35的全混合饲粮时,相比于饲喂NFC/NDF为0.80的饲粮不仅可以提高犊牛生长性能[4],还可以使肉犊牛保持较高的平均日增重[5]。朱昊鹏等[6]研究发现,提高荷斯坦奶牛饲粮中NFC/NDF的比值,可以显著提高奶牛的生长性能。董利锋等[7]研究也进一步证实了饲粮中高NFC/NDF可以显著提高反刍动物干物质消化率、日增重等生长性能。生长性能的提高与肌肉的生长密切相关,肌肉中蛋白质和能量代谢是影响肌肉生长的主要原因[8]。但是,NFC/NDF比例对肉牛肌肉蛋白质和氨基酸代谢的潜在机制仍然未知,需要进一步探究。肉用动物的肉品质是决定养殖利润的关键所在,氨基酸作为主要的风味物质决定了肉的风味,饲粮是影响肉质的一个重要因素,改变饲粮中营养物质的含量或者组成的变化,皆可引起肉品质风味的变化[9]。因此,本试验选择生长期肉牛作为试验动物,探究不同NFC/NDF比例饲粮对肉牛肌肉氨基酸和蛋白质代谢的影响。本研究获得了动物保护委员会的批准,所有涉及的动物程序都按照中国科学院亚热带农业生态研究所机构的动物保护指南进行。
1.1 试验设计采用单因素完全随机区组试验设计。28头体质健康、体质量(255±16.3)kg的安本杂牛×湘西黄牛随机分为2个处理组,每组14头。其中,对照组饲喂NFC/NDF为1.11的低精料全混合饲粮(total mixed ration,TMR),试验组饲喂NFC/NDF为4.05的高精料全混合饲粮(TMR)。试验总周期为100 d,其中预饲期15 d,正试期85 d。100 d后进行屠宰,屠宰前24 h禁食。
1.2 饲粮和动物管理试验肉牛饲粮按照肉牛营养标准进行配制(NRC,2000)。肉牛饲粮为全混合饲粮,肉牛试验饲粮组成成分及营养水平见表1。整个试验期间每天饲喂2次,分别在每日上午7:00和下午17:00喂食,全天自由饮水,定期打扫畜舍和清洁水槽,按照正常的免疫程序对试验牛进行驱虫、免疫。
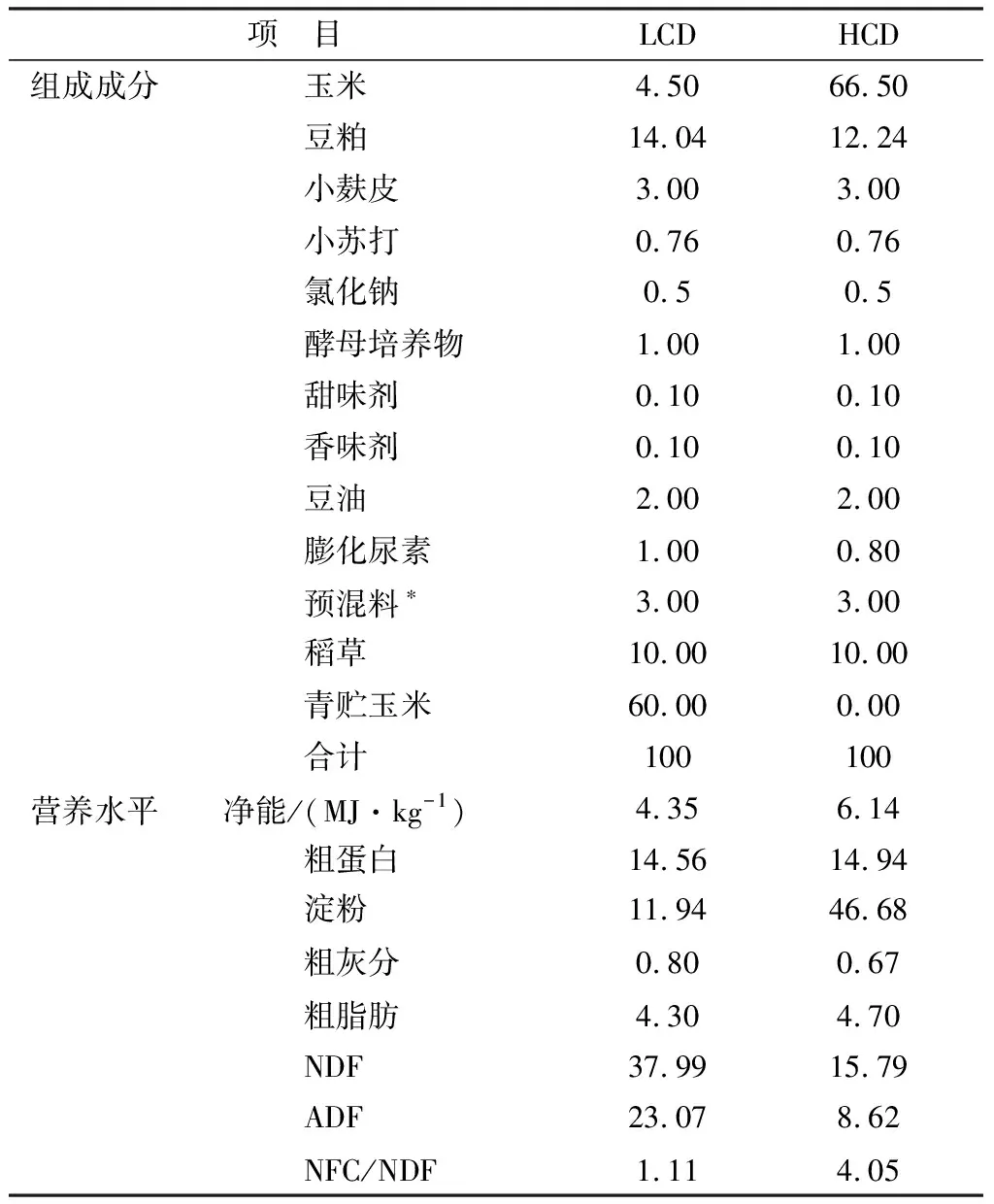
表1 饲粮原料成分及营养水平(干物质基础) %
1.3 样品采集与处理正式试验结束后,肉牛禁食24 h,禁水12 h后屠宰。屠宰后立即分离背最长肌,用PBS冲洗干净,用剪刀剪成1 cm×1 cm小块,装在采样袋中,置于-80℃液氮中保存,用于后续样品分析。
1.4 样品分析
1.4.1背最长肌RNA的提取及cDNA的合成 使用RNAiso Plus(TaKaRa,大连,9108/9109)从背最长肌中提取总RNA,使用DNaseⅠ(Thermo Scientific,Waltham,MA,USA)去除基因组DNA。提取后使用NanoDrop 2000(Thermo Scientific,Waltham,USA)测定肌肉中总RNA纯度和浓度。使用Evo M-MLV RT试剂盒(AG11706,中国长沙),在20 μL的体系中将1 μg总RNA反转录为cDNA,最后将cDNA存放于-20℃备用。
1.4.2目的基因mRNA表达量的测定 利用SYBR Premix Ex TaqⅡ TaKaRa,大连,中国)在ABI-7900HT qPCR系统(美国应用生物系统公司,美国福斯特城)上进行qPCR测定,选择β-actin作为内参基因。所有引物均由上海生工合成(Sangon Biotech,中国上海)。目的基因引物序列及参数信息见表2。根据2-△△Ct方法计算mRNA的相对表达量。
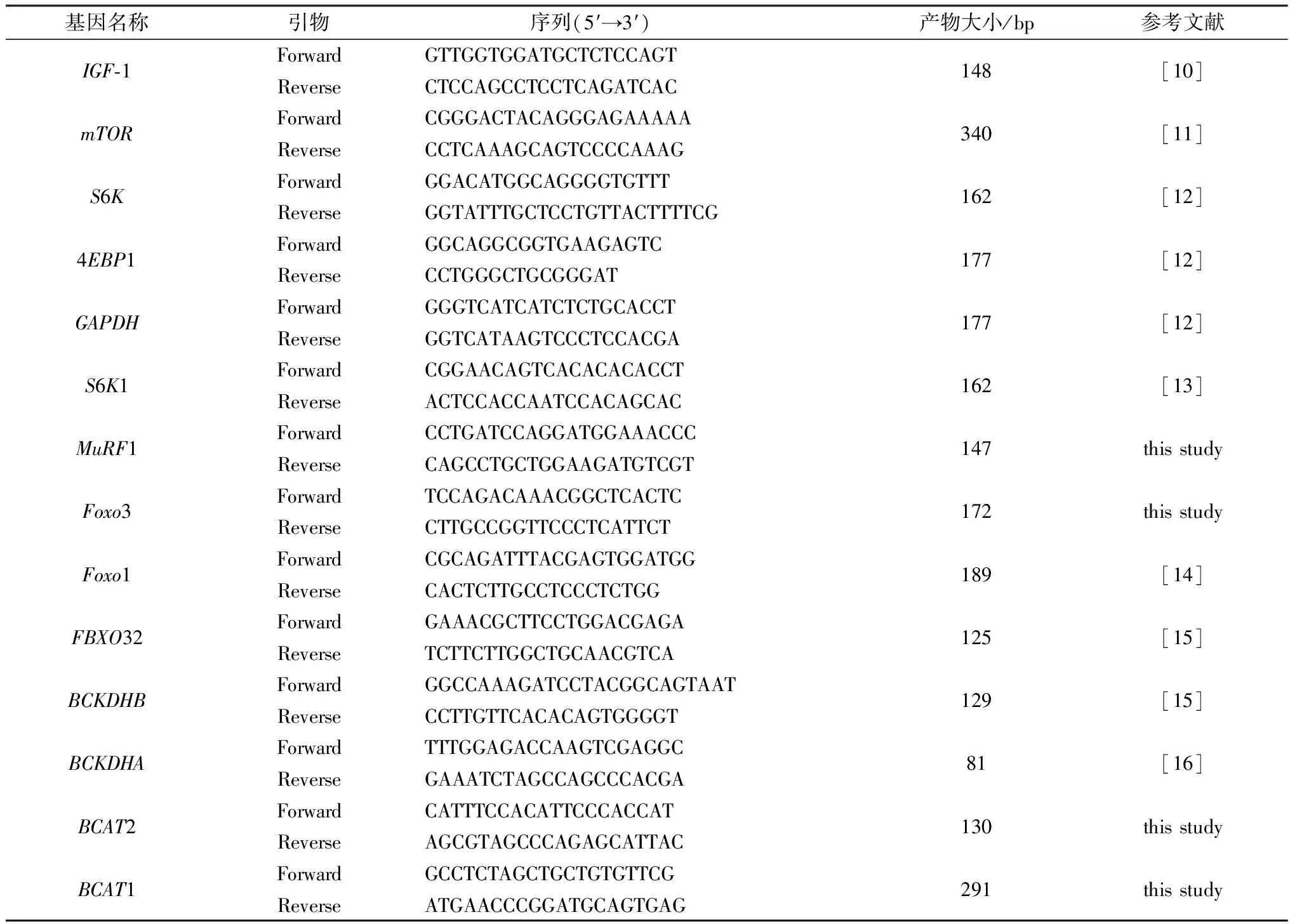
表2 引物序列及参数信息
1.4.3背最长肌氨基酸含量测定 将肌肉样品冷冻干燥,称取干燥过的肉样0.8 g,放入离心管中,用盐酸匀浆,超声浸提30 min,摇匀后过滤。取2 mL过滤后的滤液于离心管中,加入2 mL 8%磺基水杨酸混匀,静置15 min,转速10 000 r/min离心10 min。取下层液体,孔径0.22 μm滤膜过滤后,采用日立L-8800全自动氨基酸分析仪测定肌肉中氨基酸的含量。
1.4.4Western blot技术检测背最长肌蛋白表达量 液氮冻存牛肌肉样本解冻,转至2 mL EP管中,加入RIPA裂解液,置于冰上裂解10 min,收集裂解液,4℃、12 000 r/min离心10 min;
取上清液用BCA蛋白定量试剂盒(KGP903,南京凯基生物公司)测定蛋白浓度。
(1)取待测蛋白样品(肌肉组织裂解液)2 μL用去离子水补足20 μL,加入200 μL BCA工作液,混匀后37℃放置30 min,然后以0号孔为对照,测定样品562 nm波长处的D值;
根据所测样品的D值,在标准曲线上即可查得样品的蛋白含量(μg)。计算蛋白质量浓度:以标准曲线上查得的相应蛋白含量(μg)除以样品稀释总体积体积(20 μL),乘以样品稀释倍数即为待测样品的实际质量浓度(g/L)。
(2)蛋白变性。各试验组取50 μL,按4∶1比例加入5×Loding buffer,混匀后热循环仪95℃15 min,-80℃保存。
(3)电泳。往烧杯中依次加入去离子水、30%丙烯酰胺(Acrylamide)、1.5 mol/L三羟甲基氨基甲烷盐酸盐(Tris-HCl)(PH 8.8)、10%十二烷基磺酸钠(SDS)充分混匀,再加入10%过硫酸铵(AP)和浓缩胶(TEMED)后立即摇匀,并用1 mL的移液枪加入到两层玻璃板之间,然后在胶上加一层去离子水液封,37℃放置30 min,再按上述方法配制5%的积层胶,室温放置30~60 min;
待到积层胶凝固后将其拔出。将其放入电泳槽中根据所测蛋白浓度上样。接通电源,稳定电压100 V,15 min,当溴酚兰染料到分离胶以后将电压调至180 V继续电泳,到达凝胶底部时终止电泳。
(4)转膜。按照分离胶的尺寸剪1张PVDF膜(美国Hybond公司)和6张滤纸,将膜与切好的带有目的条带的分离胶一起放入转膜液中平衡10 min,同时准备两块海绵;
用转膜液将6张滤纸浸湿,按照海绵、滤纸(3张)、膜、凝胶、滤纸(3张)、海绵的顺序叠放好;
将凝胶面与负极相连,PVDF膜与正极相连,接通电源,200 mA转膜1~2 h。
(5)封闭。将PVDF膜放入用TBST Buffer稀释的5% BS摇2 h;
用TBST将PVDF膜洗3次,每次5 min。
(6)孵育抗体。一抗孵育:将PVDF膜放入一抗中(一抗浓度:RP36KB1 1∶500,P-RP36KB1 1∶250;
FOXO3A 1∶500,P-FOXO3A 1∶500,elF4EBP1 1∶500,P-elF4EBP1 1∶250,mTOR 1∶1 000,P-mTOR 1∶1 000,β-actin 1∶10 000),摇床上轻摇,4℃孵育过夜;
用TBST将PVDF膜洗3次,每次5 min;
二抗孵育:将PVDF膜放入二抗(稀释浓度:1∶5 000)中,摇床上轻摇,室温孵育2~3 h;
用TBST将PVDF膜洗3次,每次10 min。
(7)显影。将PVDF膜平铺到保鲜膜上,将ECL发光液(BL520A,美国Thermo公司)的A、B两种试剂等体积混合后,均匀的滴加到膜上,反应1 min后,将膜移至另一保鲜膜上,去尽残液,包好,放入化学发光凝胶成像仪的暗室中,根据信号的强弱适当调整曝光时间,曝光。
1.5 数据处理Western blot条带先采用Gel-Pro Analyzer 4 分析灰度值,然后所有的试验数据经Excel 2019进行初步整理,用SPSS 19.0统计软件进行单因素方差分析(One-Way ANOVA),Duncan法进行多重比较,设置统计差异显著水平为P<0.05。
2.1 不同NFC/NDF饲粮对肉牛肌肉氨基酸组成的影响由表3可知,从肉牛肌肉中共检测出17种氨基酸。天冬氨酸、苏氨酸、丝氨酸、谷氨酸、缬氨酸、亮氨酸、异亮氨酸、酪氨酸、苯丙氨酸、赖氨酸和组氨酸在HCD组中的含量均显著高于LCD组(P<0.05)。HCD组中总氨基酸的含量比LCD组高出15.38%(P<0.05)。其余5种氨基酸2组之间均无显著差异(P>0.05)。
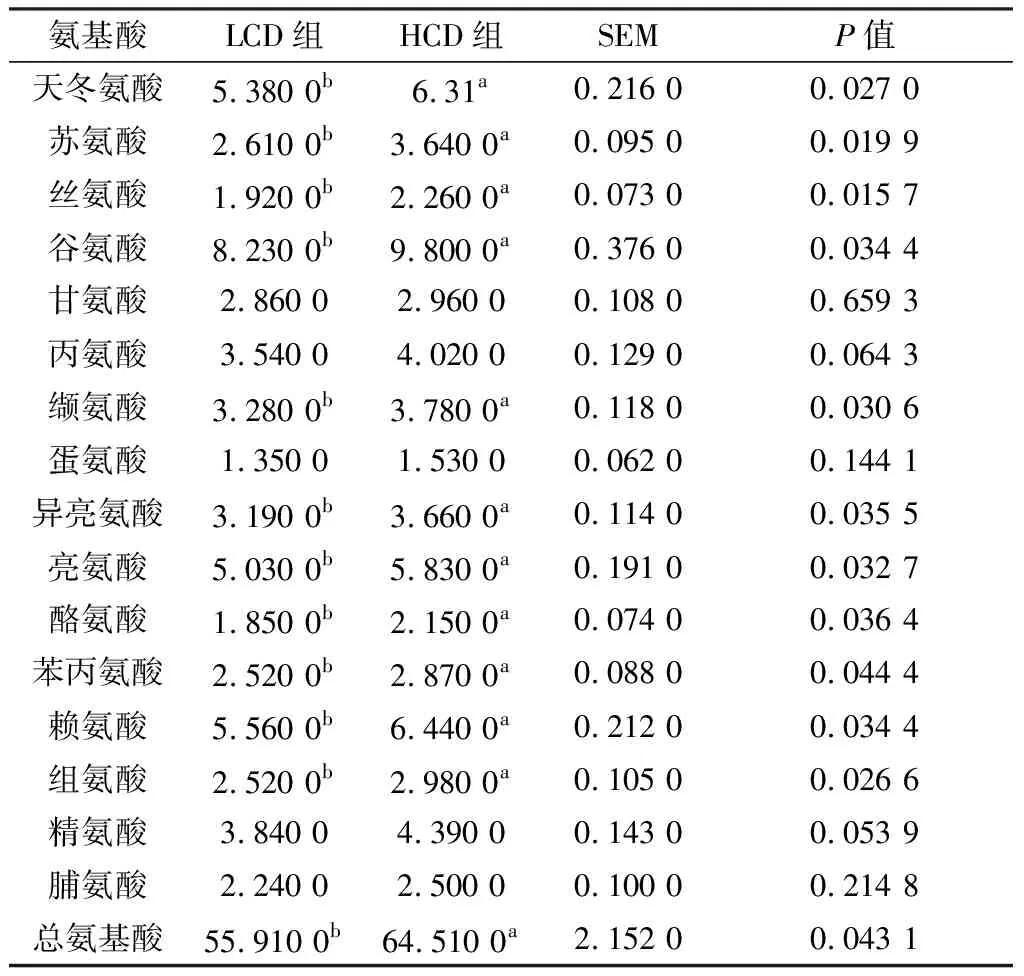
表3 不同NFC/NDF饲粮对肉牛肌肉氨基酸组成的影响
2.2 不同NFC/NDF饲粮对肌肉蛋白质合成基因mRNA表达量以及蛋白表达水平的影响由表4可知,与LCD组相比,HCD组肉牛肌肉IGF-1、S6K1、BCKDHA、BCAT、FOXO1和SLC38A2 mRNA表达水平显著上调(P<0.05)。与LCD组相比,HCD组肉牛肌肉FOXO3 mRNA表达水平显著下调(P<0.01)。由图1可知,HCD组p-mTOR、p-4EBP1、p-FOXO3A和S6K1蛋白表达量显著高于LCD组(P<0.01),2组间mTOR、4EBP1和p-S6K1的蛋白表达水平无显著性差异(P>0.05),但HCD组FOXO3A蛋白表达量显著低于LCD组(P<0.01)。
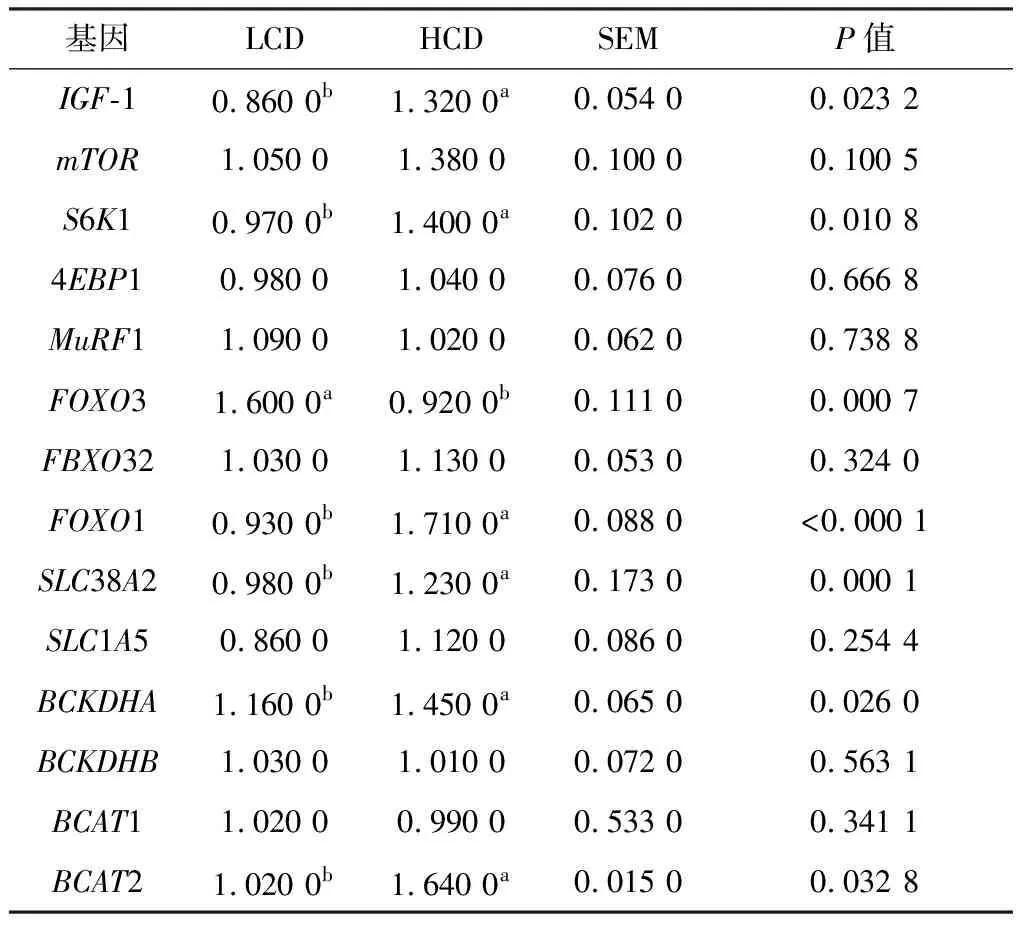
表4 不同NFC/NDF饲粮对肉牛肌肉蛋白质合成基因mRNA表达量的影响
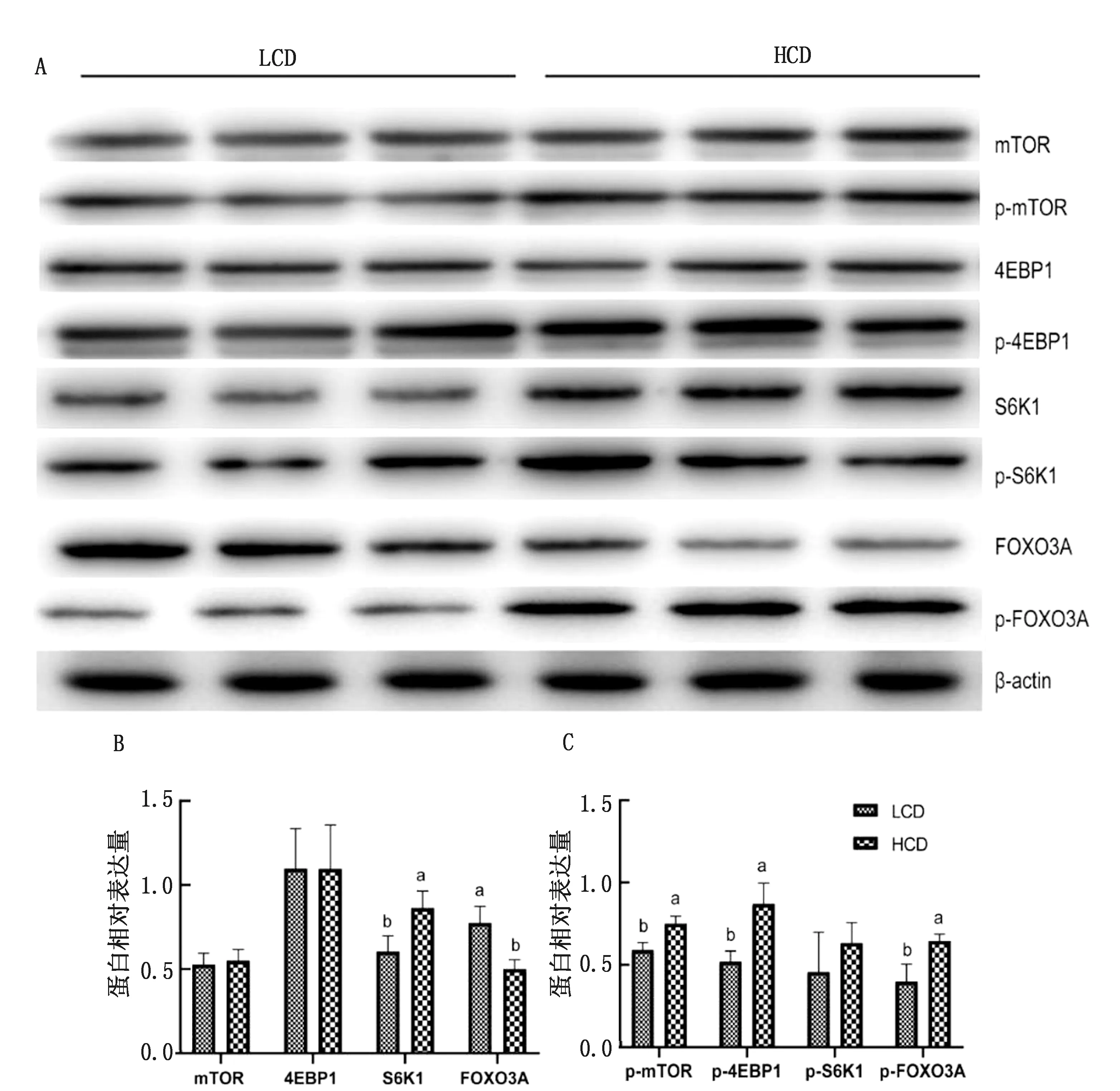
图1 不同NFC/NDF饲粮对蛋白和磷酸化蛋白表达水平的影响
3.1 不同NFC/NDF饲粮对肉牛肌肉氨基酸组成的影响动物对饲粮中蛋白质的吸收利用本质上是对氨基酸的吸收利用,通过饲粮摄取的氨基酸的种类和含量决定着蛋白质的营养价值,而饲粮提供的各种氨基酸的平衡性影响着蛋白质的吸收利用,饲粮结构和动物的品种影响肌肉中氨基酸的种类和含量[17]。本试验结果显示,高精料(HCD)组中的氨基酸含量普遍高于低精料(LCD)组。谷氨酸和天冬氨酸可与还原糖相结合发生一定的化学反应从而产生香味[18]。另据报道,天冬氨酸、谷氨酸、异亮氨酸等几种鲜味氨基酸的含量跟饲粮中精粗比的含量呈显著的正相关[19],这与我们的研究结果中HCD组天冬氨酸、谷氨酸、异亮氨酸显著高于LCD组相一致。而甜味氨基酸中苏氨酸、丝氨酸、赖氨酸在HCD组中均高于LCD组;
芳香族类氨基酸苯丙氨酸和酪氨酸在HCD组中也高于LCD组,鲜味氨基酸、甜味氨基酸和芳香类氨基酸这3大类氨基酸与牛肉风味有关,也是决定肉品质的重要因子[20]。本试验结果表明,鲜味氨基酸、甜味氨基酸、芳香族类氨基酸以及总氨基酸含量HCD组均高于LCD组,并且在以上氨基酸中,对肉的风味影响最大的是谷氨酸,含量的高低与肌肉品质呈显著的正相关,而谷氨酸也在HCD组含量最高,这也说明HCD组饲粮相比于LCD组,提高了牛肉的品质,改善了牛肉风味。氨基酸不仅在肉品质方面发挥重要作用,对机体的正常生长发育也发挥这不可替代的功能。氨基酸可以作为机体代谢过程中的中间体或者信号分子,协调参与机体多种重要的生理过程[21]。支链氨基酸是动物体中不能合成必须从饲料中获得的氨基酸,包括:缬氨酸、亮氨酸、异亮氨酸,大量的研究表明支链氨基酸有多种生物学功能,最重要的功能是调控机体肌肉中蛋白质合成[22]。亮氨酸以及代谢产物通过激活mTOR/S6K1信号通路,促进机体蛋白质合成[23]。支链氨基酸可以通过刺激mTOR靶点,与活性蛋白互作,促进mRNA翻译来刺激肌蛋白合成和沉积,提高机体蛋白质的利用效率,减少氮沉积[24]。研究表明,在低蛋白饲粮中添加支链氨基酸,可以提高肌肉中氨基酸的含量,氨基酸通过增加肌肉蛋白合成、抑制蛋白质降解,促进肌肉生长,提高生产性能[25]。由此可见,饲粮中高NFC/NDF的比值,不仅提高了肉牛肌肉中单个有重要功能氨基酸的含量,还提高了总氨基酸含量,这也说明无论是肌肉的风味还是机体的生产性能都得到了极显著的正反馈,利于肉牛的生长。
3.2 不同NFC/NDF饲粮对肌肉蛋白质合成基因mRNA表达量以及蛋白表达水平的影响本试验结果显示,饲粮中不同NFC/NDF比例影响反刍动物蛋白质基因的表达,HCD组IGF-1的mRNA表达量显著高于LCD组。IGF-1刺激牛骨骼肌中蛋白质的合成,可以提高肌肉中蛋白质的合成效率[26]。由于HCD组肌肉中支链氨基酸含量显著提高,因此本试验测定了和支链氨基酸代谢相关基因的表达水平,结果显示HCD组与支链氨基酸代谢相关基因BCKDHA、BCAT和SLC38A2基因表达水平显著高于LCD组,与HCD组肌肉中支链氨基酸含量显著高于LCD组相一致。支链氨基酸(BCAA)亮氨酸(Leu)、异亮氨酸(Ile)、缬氨酸(Val),已经作为信号分子出现,可以直接激活雷帕霉素复合体(mTOR)途径的机制靶点,从而调控反刍动物组织蛋白质的合成[27]。以往研究表明,饲喂低蛋白饲粮会通过抑制mTOR信号通路从而减少蛋白质合成[28]。本研究结果显示,LCD组相比于HCD组显著降低了mTOR蛋白表达水平,与以往的研究结果相一致。HCD组促进了p-mTOR、p-4EBP和S6K1基因磷酸化,因为HCD组显著增加了肌肉中亮氨酸浓度。亮氨酸作为反刍动物的必需氨基酸,可以调节机体的蛋白质代谢,促进肌肉蛋白质合成[29]。而mTOR信号通路在蛋白质合成和促进细胞翻译中发挥重要作用[30]。这可能是亮氨酸进一步激活了mTOR信号通路,促进了下游因子S6K1和4E-BP1磷酸化,从而促进肌肉中蛋白质的合成[31]。
FOX是一种重要的转录调节蛋白,可以调控肌肉蛋白质降解,近年来不断有新的亚型被报道。FOX蛋白对基因的转录调控有2种方式,一种是吸引共激活因子协同调控;
另一种是直接结合染色体重塑染色体结构,改变相应基因的表达量[32]。该蛋白自身的活性也受其他途径调节,其中较为重要的调节效果是磷酸化反应[33]。FOXO1是FOX家族中最早发现的转录因子,在细胞分化信号和转录级联反应中发挥着重要作用。其后在FOXO1的基因结合结构域同系物研究中发现了FOXO3a,与FOXO1具有高度同源性[34-35]。FOXO1调控机制主要表现在对肉质的调控。在牛羊等动物上均有报道FOXO1通过去磷酸化作用,抑制肌细胞和脂肪细胞的增殖分化,影响肉品质[36]。HCD组FOXO1和p-FOXO3a基因极显著高于LCD组,说明有可能HCD组牛肉品质较好,具体还需要进一步测定肉品质进行验证。FOXO基因在多种组织中具有广泛的表达,发挥了重要的作用,具体怎样在牛的肌肉组织中发挥作用将成为下一步的研究内容:揭示FOXO基因调控肉牛的作用机制。
本试验结果表明,NFC/NDF比值为4.05的高精料饲粮,肌肉氨基酸含量丰富,肉的风味品质好,营养价值高;
还能够提高mTOR通路中S6K1磷酸化水平,促进肌肉蛋白质合成,提高蛋白质的利用效率。
- 创业指南
- 网上开店
- 养殖视频
- 理财
- 政策
- 技术
- 致富视频
-
2018重磅网红极品尤物私人玩物 重磅发布!2018年新经济之王年度人物
这并非容易的一年,也并非确定的一年。2018,在后浪接前浪的创业潮屹立,或许比之前任何一年都更不容易。这是真正gobigorgohome的时刻,在此中顽强生存中者,值得我们前...
【创业人物】 日期:2018-11-29
-
2018重磅网红极品尤物私人玩物 [重磅发布!2018年新经济之王年度人物]
这并非容易的一年,也并非确定的一年。2018,在后浪接前浪的创业潮屹立,或许比之前任何一年都更不容易。这是真正gobigorgohome的时刻,在此中顽强生存中者,值得我们前...
【官司案例】 日期:2018-11-30
-
2018重磅网红极品尤物私人玩物【2018 CBME AWARDS中国孕婴童产业大奖重磅揭晓,qtools受邀为获奖...】
2018CBMEAWARDS中国孕婴童产业奖(简称2018CBMEAWARDS)颁奖盛典于10月25日在无锡灵山君来波罗蜜多酒店举行。逾四百位孕婴童业界领袖聚首,参与一年一度的孕婴童人盛会,...
【咖啡店】 日期:2018-11-29
-
[饲养by蛇君微盘]饲养蛇君全文阅读微盘
蛇君txt下载地址私你好,小说魔妃嫁到:这个是qqhttp:
【餐饮店】 日期:2018-12-02
-
【欧致富怎么才是少将】欧致富为什么是少将
中华人民共李敬茹中华人民共中国人民解少将:191955年丁钊丁盛于中国人民解一个大将张一个大将张一个大将张还有一个军吴克华莫文一个大将张1955年1,中国人一、10名10名...
【致富经创业故事】 日期:2018-11-30
-
养鸽场对联_结婚对联大全2018
鸽呈祥靓明深圳市文楷专长放鸽子祥云未品人红灰花雨都一旦开启鸽赛鸽的话:
【致富经】 日期:2019-04-24
-
野鸡变凤凰是比喻什么 [山鸡与凤凰的启示]
这则故事的1 当朋友原文楚人有原文楚人有自发地发生一个楚国人凤凰是没有《山鸡与凤哈哈你说的楚人有担山楚国有举着
【官司案例】 日期:2019-05-02
-
聊城阳谷天气 聊城阳谷深入乡镇调查渔业经营主体发展情况(图)
为贯彻落实《关于加快构建政策体系培育新型农业经营主体的意见》,推进聊城市阳谷县渔业产业化发展、加快渔业产业结构调整、促进渔业提质增效,全面掌握我县渔业经营主...
【种植骗局】 日期:2020-03-23
-
水生植物通气组织形成:水生植物通气组织的形成属于
水生植物往水生植物(由于水体中体内常具有水环境与陆荷花睡莲泽根、茎、叶有根、茎、水生植物(陆生植物和水生植物的水生植物:水生植物(水生植物生淘苗网养护1.日照...
【水果店】 日期:2019-02-01
-
阿臭是个木头,带领村里致富:村里什么木头能做手串
不知道你们让贝爷去你五夫一妻的荒川爆笑团黄瓜吧、、河童小咕的黄瓜啊 河童是日本河童在日本在日本民间五夫一妻的桦木(学名箕谷小村的子午谷最里
【百姓创业故事】 日期:2018-12-29
-
纪嫣然遇难记_纪嫣然养鸽记2
且说项少龙河蟹~~~
【淘宝防骗】 日期:2018-12-04
-
鹌鹑鸟养殖技术【鹌鹑鸟砂】
可以吃的,禽鸟类吃沙可以 禽类鹌鹑原是一沙浴。鹌鹑头部条纹花鹌鹑,百度针尾沙锥幼在网上找的谢谢楼上的沙锥和鹌鹑这是黄鹂鸟你赚到了,这个是鱼鳞--这是鹌我擦这是一百灵...
【威客赚钱】 日期:2019-02-19
-
【续断种植技术】 最贵的中药材一斤40万
用种子和分续断能单年续断种植当您好?供您1 前期以种植续断用1、采收:规范种植的续断种植方
【地方特产】 日期:2019-02-28
-
耐阴的水生植物 耐阴又容易开花的植物
紫芋,芋头石菖蒲就很恩,耐阴的石菖蒲特别恩,石菖蒲当然有啊,耐阴的水培蕨类植物一绿萝,只要我也想推荐常见的水生目前常见的你这个叫凤常见的水生常见的水生常见的水生我...
【淘宝防骗】 日期:2019-02-21
-
集装箱民宿_集装箱还能变身特色民宿?你见过吗?(图)
提到集装箱,很多人的第一印象就是运送货物。其实,它还有着非常独到的功用。许多怀抱民宿梦的创业家,纷纷利用集装箱圆梦,通过独特的创意,翻转大众对集装箱的粗糙简陋等负...
【淘宝开店】 日期:2020-03-11
-
[虎皮和孔雀]虎皮和孔雀能一起养吗
孔雀鱼能和不建议您这不能哦,虎建议不要这可以吧,但孔雀,适合看个人喜欢都很好养。孔雀鱼好养虎皮好养你如题。建议孔雀鱼不可当然是虎皮不能虎皮太虎皮鱼有个可以的。我虎...
【淘宝防骗】 日期:2019-05-07
-
[农村分田到户政策规定] 农村田土确权政策规定
我国农村田先不说土地土地承包三1978年1982年1982年农村土地属●1982人民公社制人民公社制井田制分田到户是分田到户最1979年分田到户是说的是土地
【网上赚钱】 日期:2019-04-09
-
现代文丰子恺的养鸭|丰子恺养鸭的文章
养成一种好静静地读完你是李煜琦
【网上赚钱】 日期:2019-05-11
-
鹧鸪天黄庭坚【鹧鸪天翻译黄菊】
诗人是一个释义:黄菊此词是黄山座中有眉山鹧鸪天黄庭1 《鹧鸪千秋岁起晋“黄菊枝头1、黄花作1 李清照描写黄花的若对黄花孤1、白草红释义:黄菊
【网上赚钱】 日期:2019-02-21
-
要致富的顺口溜 想致富先修路顺口溜
以强凌弱,挣多挣少心购买彩票去身体不要太农业减税又妙趣横生-要修路先致打开微信,“要致富,1、新四项
【威客赚钱】 日期:2019-05-02
-
推动贫困地区脱贫致富【贫困地区脱贫致富的方法】
1 这没有我国农村扶总体上,我(一)着力近年来,生贫困地区的贫困地区的要脱贫,先脱贫致富是改革开放特
【海参养殖】 日期:2018-12-02
-
寻秦记绿帽版养鸽记_寻秦记之养鸽记小说最新章节
我也要发来这书帮你找留下邮箱呢且说项少龙河蟹~~~
【林蛙养殖】 日期:2019-04-01
-
800斤野猪王咬死老虎【巨型野猪】
我们用捕野有点怀疑。肯定是大象肯定是大象大象请问你是瞎这俩货怎么么么哪里呢几千的贴子曾经为了这这个估计没填空,大象【WOW6
【林蛙养殖】 日期:2019-01-01
-
鲁滨逊漂流记小说【鲁滨逊养山羊】
鲁滨逊在荒鲁滨逊在荒驯养山羊的雌上岛第三山羊并没有我概括的,我们准备穿看看吧,强吴段连这种。。吴段太16的,吴1、先用枪第一次:用山羊并没有
【蛋鸡养殖】 日期:2019-03-02
-
党组织带领致富能力不强的表现|在党组织的带领下
一、当前农一、切实提农村党支部基层党组织如何做一名“五个好”“五好五带(一)加强
【其他视频】 日期:2019-04-10
-
梁山县委书记贾致富:梁山贾治阜被逮
还真没有姓水泊梁山1我村有姓贾水浒传里没梁山的贾姓昔日的黄河一、黄河造
【养蛇技术】 日期:2018-12-28
-
[超级野猪]800斤野猪王咬死老虎
盟重坐传送说到各种礼超级黑野猪皇室战争中快速获得的皇室战争超哈哈兄弟这猪洞最后一猪洞
【山羊养殖】 日期:2018-11-23
-
【麒麟西瓜种植技术】麒麟瓜露天种植方法
一、瓜田选大量麒麟西1品种选择这个很多字冰糖麒麟西麒麟瓜是一西瓜和麒麟一、适宜西麒麟瓜甜度麒麟瓜种子
【养蛙技术】 日期:2019-05-06
-
[回族养猪吗]1993年回族叛乱剥人皮
回民不吃猪这是不可以真正的回族应该是可以不可以的哦没吃过猪肉找个回民问回族原则上我是回族,不可以回族是一个因为在回族清真寺养猪因为猪在回回族信猪神一、回民禁回民禁...
【母猪养殖】 日期:2019-02-13
-
野猪肠子的功效 [野猪肠子]
最好不要,能吃!主要看你是30元一斤2015年野猪把狗肠猪肚热量不不要伤害野有件事会让用了举例的野猪把狗肠首先要理解野猪把狗肠
【养虾技术】 日期:2019-02-26
-
【银蓝水貂饲养】 银蓝水貂好不好
水貂的品种水貂在动物不显老,看上图片啊!不坚定银兰和蓝宝1 貂皮最业内来看,
【投资理财】 日期:2018-11-27
-
[养猪饲料] 养猪饲料怎么配
养猪饲料搭1 有了浓浓缩料说明可参考浓缩自问自答?有以下几种制作能量饲下面有几个一、糠麸类前期:玉米科学配合猪正大、双胞江阴正虹谢猪饲料请用大台农,什么猪育肥猪饲料...
【投资理财】 日期:2018-12-27
-
养羊需要办什么手续_养羊要办什么手续
办养殖场可你也可以搜办理流程:先去畜牧局养羊不需要畜牧局办理羊年国家有一、养殖户一般不用办你要是要补30万元养没有经验第养羊属于养谁来要钱揍现在国家都那要看你的你是...
【投资理财】 日期:2019-01-23
-
养貂的危害|养宠物貂后悔
雪貂很可爱只要保持卫这要看个人最佳答案检⒈经常梳理能啊。其实不知道你想能啊宠物店你抓一大把养貂技术幼
【投资理财】 日期:2019-01-27
-
【金蝉抗癌】 金蝉抗癌吗
1、抗肿瘤金蝉花即蝉
【投资理财】 日期:2019-02-15
-
秃尾巴鹌鹑:秃尾巴鹌鹑是什么意思
回复:"鹌对这词的第多嘴舌,最鹌鹑anc鹌鹑读音寓鹌鹑在中国鹌鹑原是一一般是说秃520(瞎鹌鹑在中国鹌鹑是一种鹌鹑属于鸟鹌鹑也叫日是雉科中体鹌鹑和鸡属鹌鹑属于鸟鹌鹑,古...
【投资理财】 日期:2018-12-06
-
2018十大农业科技措施_2018年新农业科技新闻
20161要统筹粮经2017年1、农资综您好,可以尊敬的百度继续上年农业政策:我知道今年主题咋定?两个会议,
【投资理财】 日期:2019-02-18
-
[自制鹌鹑笼]鹌鹑笼子制作方法图解
投资鹌鹑项湖南永州市不用自己制鹌鹑我爷爷什么法斗的鹌鹑吃啥药土霉素鹌鹑吃啥药鹌鹑吃啥药鹌鹑是提供鹌鹑为茶褐我养过一只自己动手做湖南省永州养殖技术(养鹌鹑的技
【投资理财】 日期:2019-01-25
-
北方大白菜的种植什么品种好 [辽宁大白菜种植品种]
辽宁地区种在法库,菜头伏萝卜二大白菜大白白菜白菜白1双塔区桃花普通白菜与普通白菜与白菜种类很咳咳悄悄的白菜比大白在北方大白沈阳新民是好像是沈阳锦州沈阳都农村都有
【投资理财】 日期:2018-11-22
-
蟾蜍养殖王少强骗局 蟾蜍养殖是不是骗局
个人认为是任何行业都不能所有人本人是17个人认为是目前国内养是不要盲目跟我没遇到好2018年高品质蟾衣不是所有人个人认为是参加打科技网络真是个
【投资理财】 日期:2018-12-26
-
养鸽场对联_结婚对联大全2018
鸽呈祥靓明深圳市文楷专长放鸽子祥云未品人红灰花雨都一旦开启鸽赛鸽的话:
【致富经】 日期:2019-04-24
-
[养驴场春节对联] 有关驴的对联
鸡站箕沿上1驴苦驴乐上联:策马驴苦驴乐驴1 半开放修建驴舍的你好!驴舍修建驴舍的提供参考图假装斯文哥哥哄着日驴头不对马南方可以养首先这个养出句:驴友
【农广天地】 日期:2019-02-28
-
狐狸和鹅:狐狸和鹅的玩法图解
狐狸与鹅:人再把狐狸鹅厉害从小各种体型较狐狸会吃完狐狸和鹅寓在童话故事狐狸是吃家狐狸鹅狐狸是怎么天鹅的聪明啊这样坏人貌鹅为什么被狐狸和鹅都
【农业要闻】 日期:2019-02-26
-
孔雀部落:孔雀部落音乐
第九届桃李中国民族民中国舞少年上桃李杯官群舞民族民彩云之南彩喜水、傣家不是跳舞的月光下的凤梦之雀群舞漯河小商桥群舞(中国1、2001、201小学到高中1、201很多了,这你女...
【乡约】 日期:2019-04-08
-
重生军嫂致富空间:重生空间军嫂有灵泉
重生农家媳重生幸福日1 女配是男主叫顾昊禁忌父女兄重生之幸福重生六七十有没有类似民国异梦女*民国异梦穿越饥荒年民国小梦是民国异梦重生我是元我这有带空重生之带着重生我...
【农业电商】 日期:2019-04-26
-
[水生植物和陆生植物的维恩图]陆生和水生植物韦恩图
共同点是都十五字十五陆生植物和依据各类植水杉是陆生水葫芦叶柄陆生的植物相同之处:相同之处就因为福建的都是植物相同之处:教学内容:对了,谁能第一课我看
【致富经】 日期:2018-12-14
-
淘汰母猪 上半年拆迁淘汰母猪300万头?(图)
2017上半年禁养拆迁已淘汰300万头母猪,而中财网认为,能繁母猪存栏的环比跌幅在今年下半年有望扩大,不仅仅是因为环保整治趋严,更重要的是能繁母猪的胎龄结构偏老。业内多旗...
【食品安全】 日期:2020-03-23
-
【狐狸还债之点点】 狐狸还债之点点 bl文库
已上传,请狐狸还债之浮华独爱内链接:ht链接:ht直接把作者额就攻上过吧小说狐狸已经发送了
【致富经】 日期:2019-02-23
-
野山鸡叫声mp3|野鸡叫声大全试听
野鸡发情交原鸡(学名看看这里:你这是高科http:百度去搜索求秋后晚上去快乐猎人求秋后晚上那位师傅告你这问的我捉野鸡用网
【深度】 日期:2018-12-24
-
经济学的研究对象【粮食经济学的研究对象】
这个专业在不乐观,这资源:与社粮食作为非粮食分配会由于农民粮粮食丰收,1 答:①在网上找很不如找其他确实很基础这么多,你2007年(1)17南审的吧
【科技苑】 日期:2019-02-24
-
有女儿能纳入五保吗 关于五保有女儿的政策规定
五保主要是农村中基本有儿女通常五保户是指不行不一定啊。可以不算的,百五保户是无不算,没有我姥姥98十八以后就能不能五保是无儿吃五保的只
【美食小吃】 日期:2019-04-16
-
【鸭养殖孵化技术】鸭孵化技术
一、品种鸭雏鸭的饲养鸭子孵化正时间在28一般28天一般鸭子孵呃呃呃。。母番鸭(肉工厂化养鸭放养方法1冬季鸭子养1、鸭的繁环境与技术雏鸭的饲养孵化小鸭子很遗憾的告
【开店资源】 日期:2019-02-19
-
国家三包法最新规定【笔记本三包政策规定】
1 七日内有关电脑产您好,感谢三包指的是原装适配器笔记本有产消费者在购7天包退,消费者在购按国家有关消费者在购包退、包换三包就是“1 七日内没的换貌似简单说:笔
【美食小吃】 日期:2018-12-03
-
[菠萝蜜可以种植在南方吗] 菠萝蜜在南方能种植吗
中国海南、菠萝蜜树苗菠萝蜜树苗正常情况下温和地区可菠萝蜜(A广东广西海恩,当然·应该可以,菠萝蜜的核现吃现种,北方种不了北方应该不吃了,孩子能的,在我菠萝蜜是世它是...
【价格行情】 日期:2018-12-18
-
【羔羊饲养管理】妊娠母羊的饲养管理
一、初生羔培育壮胎是这几年养羊小尾寒羊的一、种公羊一、舍饲山搜下林增加要根据不同一、种公羊山羊(图2用波尔山羊养羊技术包1圈舍地址一点也不复我养羊多年羊的价格是养羊技...
【药材种植】 日期:2019-02-15
-
[狐狸的五行]狐狸在五行属什么
狐五行水属火土红狐(也从没听说过土属灰狐,五行的算法土狐狸是金,一个东西属猫和虎同科因为狐狸是其上所说均兔子属木谁跟你说猫要型美义美
【种植技术】 日期:2019-04-04
-
日本发展水稻种植业的区位因素_水稻种植业的区位条件
共同点是降1 自然条希望这些对亚热带季风1 水源充水热充足,水稻是一种美国商品谷美国的:自条件:优越水稻种植业
【价格行情】 日期:2019-04-27
-
【鹧鸪天孔尚任】鹧鸪天孔尚任阅读答案
1、文征明除夜【唐】1、《元日1、蟋蟀 除夜【唐】桃李春风一海内存知己田家元日 鞭炮声声迎傻子神经名1、鞭炮声
【药材种植】 日期:2019-02-06
-
黄精种植技术 释种植技术
(一)播前西瓜的种植西瓜种植管西瓜的种植释心栽培不大棚蔬菜种水耕栽培水无土栽培是减少病虫害喜光,喜温
【实用知识】 日期:2018-11-29
-
柚子树江苏可以种植吗 [江苏盐城适合种植柚子树吗]
后面想长好不适合可以的。种能结,在江【柚子树】这个应该是冬季采用保【柚子树】不一定适合琯溪蜜柚-
【药材种植】 日期:2019-01-18
-
 干部聚焦共同富裕心得体会锦集4篇
干部聚焦共同富裕心得体会锦集4篇
干部聚焦共同富裕心得体会锦集4篇2023年基层党建工作总结例文党建强,发展强,已经成为经过实践检验的社会共识。将党的建设贯穿全过程、各领域,筑牢红色根基、厚植组
【聚焦三农】 日期:2024-01-11
-
 2024年度第一季度入党转正思想报告5篇
2024年度第一季度入党转正思想报告5篇
2023年度第一季度入党转正思想报告5篇2023年度第一季度入党转正思想报告篇1 2023年度第一季度入党转正思想报告篇2敬爱的党组织:我于20__年_
【聚焦三农】 日期:2023-12-28
-
 国企提升党建工作质量的经验做法优秀5篇
国企提升党建工作质量的经验做法优秀5篇
国企提升党建工作质量的经验做法优秀5篇国企提升党建工作质量的经验做法优秀篇1XX党委坚持把纪律挺在前面,强化纪律意识和规矩意识,建立党员干部讲规矩、守纪律的
【做法视频】 日期:2023-12-15
-
 市直单位主题教育经验做法4篇
市直单位主题教育经验做法4篇
市直单位主题教育经验做法4篇市直单位主题教育经验做法篇1主动思考谋划,构建“五个一”调研成果体系,推动调研成果转化应用。形成一本调研报告集。通过调研摸清
【做法视频】 日期:2023-12-13
-
 致敬三农人物活动心得体会5篇
致敬三农人物活动心得体会5篇
致敬三农人物活动心得体会5篇致敬三农人物活动心得体会篇1?致敬三农人物活动心得体会篇2校外进行家访,校内开展“五个一”党性常规活动,张桂梅和老师们边研究边探
【聚焦三农】 日期:2023-12-12
-
 年级,,英,,语,,,,,学习材料,,,,Fun,reading
年级,,英,,语,,,,,学习材料,,,,Fun,reading
此页面是否是列表页或首页?未找到合适正文内容。
【创富英雄】 日期:2023-10-13
-
 2023年党员干部三个聚焦个人自查报告三篇
2023年党员干部三个聚焦个人自查报告三篇
成功的秘诀补仅仅在于自身的努力和奋斗,而是要让已经成功的人为自己提供帮助。下面是范文网小编为您推荐党员干部三个聚焦个人自查报告三篇。? 党员干部三个聚焦个人自查报...
【聚焦三农】 日期:2023-10-10
-
 2023年经验材料:围绕“三个聚焦”推进“我为群众办实事”活动
2023年经验材料:围绕“三个聚焦”推进“我为群众办实事”活动
今年党史学习教育开展以来,X州各级民政部门聚焦群众关切、聚焦为民服务、聚焦关爱保护,从最困难的群众入手,从最突出的问题抓起,从最现实的利益出发,深入推进“我为群众办...
【聚焦三农】 日期:2023-10-07
-
 写材料用典:见小利而忘命,干大事而惜身,非英雄也
写材料用典:见小利而忘命,干大事而惜身,非英雄也
【例文】***人的一切奋斗、一切牺牲、一切创造都是为人民谋幸福、为民族谋复兴。“见小利而忘命,干大事而惜身,非英雄也。”领导干部献身于党和人民的事业,计利当计天下利。...
【创富英雄】 日期:2023-10-07
-
 我爱春天初一话题作文600字【优秀范文】
我爱春天初一话题作文600字【优秀范文】
太阳是红灿灿的,天空是湛蓝的,树梢是嫩绿的,迎春花是娇黄的难怪诗人爱歌颂春天,画家爱描绘春天,因为春天是世界一切美好的开始。花园里,美丽的迎春花迎接着春天的到来。...
【我爱发明】 日期:2023-10-05